ADAPTIVE VERSUS MALADAPTIVE EMOTIONAL TENSIONVadim S Rotenberg vadir@post.tau.ac.il
Wolfram Boucsein
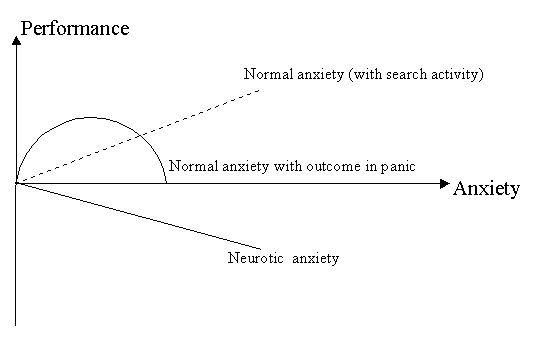
Genetic., Soc ml, and General Psychology Monograph, 1993, 119(2), 207-232. Received July 23, 1992 ABSTRACT The problem of emotional tension is usually discussed only with regard to its negative (maladaptive) aspects The positive (adaptive) function of emotional tension, however, is as important as its negative function In this article, we have examined psycho physiological outcomes of adaptive versus maladaptive emotional tension with respect to opposite forms of behavior (search activity and renunciation of search) that have an opposite outcome on body resistance and performance Some traditional psychophysiological problems, for example, the law of initial values (Wilder, 1931), the difference between orienting and defensive reaction, or between anxiety and panic behavior, are revised on the basis of the assumption of adaptive and maladaptive emotional tension. Emotional tension plays a significant role in various contexts, although it is most frequently discussed and investigated with respect to its negative aspects, which are subjectively experienced as distress, fear, or pathological anxiety Emotional tension also has positive effects, enabling an individual to overcome various obstacles, and thus plays a positive role in the adaptive process Emotional tension, therefore, can be either maladaptive or adaptive in both nature and outcome Psychobiologically, there are two main requirements for the process of adaption (a) It should enable the individual to continue his or her goal-directed behavior, and (b) it should minimize physiological costs, meaning that body resistance must not decrease. The subjective emotional experience in the process of goal-oriented behavior may vary from joyful anticipation and pleasure to anxiety and anger Nevertheless, as long as the goal-oriented behavior continues to occur and the body resistance is not decreased, even very unpleasant emotional experiences cannot be regarded as maladaptive. In other words, the difference between adaptive and maladaptive emotional tension is not the same as the difference between positive and negative emotional experiences. For example, negative emotional tension predominated in German concentration camp prisoners during World War II, but they continued their everyday struggle for survival and survived (Chodoff, 1970). And the absence of negative emotions or even a predomination of the positive emotion of satisfaction does not protect against the so-called diseases of achievement (psychic or psychosomatic disturbances that sometimes appear after the goal is achieved, but reflect maladaptation). Despite the theoretical and practical importance of the distinction between adaptive and maladaptive emotional tension, neither adequate theoretical approaches nor valid methods appear in past research to distinguish between the two. Our first aim in this article is to give some theoretical background for such a twofold concept and to support it by behavioral and psychophysiological data taken from human and animal studies. An integrative neurophysiological model is also provided, focusing on the relation of brain monoamines and different kinds of arousal. Finally, the implications of a psychophysiological approach that differentiates between adaptive and maladaptive emotional tension are discussed, including endocrine measures. Theoretical Approaches to Different Forms of Emotional Tension Emotional tension may be embedded in different theoretical contexts. In an approach closely related to general activation theory, emotional tension is regarded as a qualitatively unitary state that only varies quantitatively. According to this approach, an optimal level of emotional tension is adaptive in helping the individual to solve problems and to overcome obstacles without any negative outcome for the organism, whereas according to the Yerkes-Dodson law (Atkinson, 1974; Lens & DeVoider, 1980; Yerkes & Dodson, 1908), if the level of tension is extremely low or high, its outcome is regarded as negative with respect to performance, adaptation, and health. Nevertheless, there are many exceptions to the Yerkes-Dodson law. First, pathological emotional tension is always harmful, with respect not only to health but also to performance, thereby decreasing the effectiveness of the latter in a linear manner (Lader, 1975; Ramirez & Lasater, 1976; Spielberger, 1962). As illustrated in Figure 1, the influence of pathological anxiety on behavioral outcome will always be negative, even in the case of low emotional tension (shown by the solid line in Figure 1). Another contradiction to the Yerkes-Dodson law is that even a very high level of emotional tension can promote the activity of the subject. According to Dienstbier (1989), naturally evoked peripheral catecholamines never seem to be too high for optimal performance. In addition, when methods of coping were available, even very high arousal levels failed to elicit discomfort and negative emotions (Gal & Lazarus, 1975). Another approach to opposite behavioral and psychophysiological outcomes is based on stimulus characteristics. The orienting reaction (OR), which is tied to exploratory behavior, as opposed to the defensive reaction (DR), which is connected with avoidance behavior, also may be differentiated by their physiological components (Graham, 1979; Lacey, 1967; Sokolov, 1963). The discrimination between OR and DR, however, is not free from ambiguities. First, OR is not opposite to DR in the sense of coping, because they both can increase body resistance. In addition, stimuli normally eliciting an OR may sometimes produce a DR instead, and vice versa. Furthermore, DR is typically considered to be a unitary reaction, although some data revealed autonomic responses in cases of avoidable and unavoidable punishment (Obrist et al., 1978) and during active coping (Harburg, Blakelock, & Roeper, 1979; James, Hartnett, & Kalsbeek, 1983).
FIGURE 1: Connections between different levels of emotional tension and outcome in normal and pathological subjects. available, even very high arousal level At first glance, the increase of heart rate (HR) and cardiac output during a DR seems to correspond to the increased metabolic requirements of the reaction's motor component, an increased metabolic rate could help to discriminate DR from the physiological reaction to the intellectual component of the task However, only during the cold pressor test and physical exercise was an exact correspondence between cardiac output and metabolic demands observable (dinner, Bedi, & Horvath, 1979, Lovallo, Pincomb, & Wilson, 1986a, Obrist, Light, McCubbin, Hutcheson, & Hoffer, 1979), although cardiac-metabolic dissociation occurred during problem solving (Carrol, Turner, & Hellawell, 1986) The same metabolically unwarranted and excessive rise of cardiac output and HR was also registered in a shock avoidance task (Sherwood, Alien, & Obnst, 1986, Stoney, Langer, Sutterer, & Gelling, 1987) When regarding physiological correlates of emotional tension, it is important to take into account qualitative physiological differences between certain kinds of behavior This requires outward-directed attention, sensory rejection, and inward-oriented attention (e g, Diamond & Carver, 1980, Lacey & Lacey, 1970) The physiological response that is adaptive for one kind of task may be inadequate for another kind, indicating unproductive emotional tension instead Because of the ambiguities and contradictions of these approaches, Rotenberg and Arshavsky (1979b) formulated another approach, inspired by the necessity to distinguish adaptive and maladaptive emotional tension as two qualitatively different psycho-biological states of the organism Adaptive emotional tension can appear only if the so-called state of search activity is predominant (Rotenberg, 1984, Rotenberg & Arshavsky, 1979b) Search activity is defined as activity that is oriented to change the situation (or at least the subject's attitude toward it) in the absence of a precise prediction of the outcome of such activity, but with respect to possible outcomes at all stages of processing Search activity can be regarded as a psychobiological state that facilitates active self-stimulation in animals, creative behavior m humans, as well as outward oriented, exploratory, and active defense (avoidance as well as fight/flight) behavior in both species (Table 1). Maladaptive emotional tension, on the other hand, appears in a so-called state of renunciation of search, which is present in pathological anxiety and depression in humans, in freezing and imaginary death in animals, and in panic and stereotyped behavior in both species These two different forms of behavior also have an opposite influence on sleep structure.
Rotenberg (1984) and Rotenberg and Arshavsky (1979b) proposed the concepts of search activity and renunciation of search to explain the complicated and contradictory results of investigations of the bodily outcome of different types of behavior One of these behaviors is fight or flight as active self-stimulation that increases body resistance to various forms of artificial pathology (e g, experimental epilepsy, peptic ulcer anaphylactoid shock, blood hypertension, myocardial infarction, cardiac rhythm disturbances, Parkinson-like neuroleptic syndrome, Rotenberg & Arshavsky, 1979b, Vainstein & Simonov, 1979, Valdman, Kozlovskaya, & Medvedev, 1979, Weiss, 1977) In contrast, freezing, imaginary death, and even passive self-stimulation decreased body resistance and accelerated the development of artificial pathology (Anderson & Jingling, 1978, Rotenberg & Arshavsky 1979b, Seligman, 1975, Vainstein & Simonov, 1979. It was therefore necessary to identify two opposite common factors that determine these two groups of different behavioral patterns and their opposite effects on body resistance and on the course of the artificial patholog. It was not the difference been positive and negative emotions, because passive and active self-stimulation both represent the positive emotional state, and fight, flight, and freezing represent the negative emotional state Moreover, different forms of behavior in each group represented separate emotional patterns (Panksepp, 1982) For instance, active self-stimulation represents expectancy, fight represents rage, and flight represents fear It was not the difference between approach and avoidance, because fight and flight belong to one group, and passive self-stimulation and freezing to another. It was not the difference between general activity and passivity, because freezing was sometimes displaced by motor irritation and panicky behavior, and it did not change the outcome on bodily resistance Moreover, when animals are stimulated to inhibit motor activity arbitrarily in order to avoid an electric shock, the successful learners had fewer lesions on their gastrointestinal tracts (Goesling, Buchholz, & Carreira, 1974) than unsuccessful learners On the other hand, when an animal is exposed to an unavoidable electric shock, ulceration of the gastrointestinal tract increased in tandem with motor activity (Weiss, 1977) Consequently, it was not the difference between activity of the sympathetic and para-sympathetic systems, the sympathetic system dominated in all forms of behavior that prevent pathology, but its activity was also prominent during freezing (Anderson & Jingling, 1978, Rotenberg & Alexeyev, 1981, Rotenberg & Arshavsky, 1979b, Valdman et al, 1979) After analyzing all of the above-mentioned factors, Rotenberg (1984) and Rotenberg and Arshavsky (1979b) concluded that it is search activity and renunciation of search that determine the course of somatic pathology Behavior of the fight-flight type is classified as search behavior because an attempt is made to overcome or avoid a stress-producing situation in the absence of a priori confidence in the success of such an attempt Self-stimulation, to some extent, also acquires features of the search for positive emotional experience in the absence of a priori confidence in the success caused by the negative emotional component of self-stimulation (Grastyan, 1976) The duration of the self-stimulation period correlates with the distinctness of the animal's general exploratory reaction (Schiff, Rusak, & Block, 1971), and such an activity exhibits a competitive relationship with stereotyped behavior (Wauquier, 1980) Self-stimulation is often produced from the same brain regions as exploratory and investigative behavior, which obviously display search activity (Panksepp, 1982). It is crucial to establish a biological basis of search activity Most probably, search should emerge in a situation in which some individual requirements cannot be satisfied by previous well-developed behavioral habits (i e, during conditions of threat or challenge) However, it would be incorrect to assume that search activity in all cases is merely a "servant" of some irrelevant requirements, even if they cover an extremely wide range, from the biological to the obviously human such as self-esteem If the organism only follows the course of a continuously changing world with the aim to adapt to it, it may lose time and initiative We assume that several independent anti-homeostatic requirements (e g., new information, new emotional experiences) can be regarded as concrete manifestations of a more general notion-a search requirement Exploratory activity without any pragmatic necessity reflects a high level of search requirement, as does creativity in humans. It is important to emphasize that the positive outcome of search activity on body resistance and adaptation is determined by the process of searching, not by its result, which is the benefit of successful search behavior It is a crucial point of the very concept of search activity, as opposed to the concept of coping behavior, which must be successful Search activity may be unsuccessful and may not help to overcome obstacles, but nevertheless, its continuation has a positive outcome on body resistance Seligman (1982) confirmed in experiments that if an animal remains active in the case of inescapable stressors, its resistance is higher than in the case of passive behavior-even in an objectively controlled situation. Although the presence of search activity is as essential to an individual as to the entire population, the intensity of search requirement may fluctuate within a very broad range Search activity can be suppressed, strengthened, or distorted Only the prerequisites for its development are genetic, whereas the realization of these prerequisites largely depends on the appropriate conditions and specific features of rearing, especially in early childhood Two opposite situations appear to be equally unfavorable for the development of search activity and search requirement conditions in which all actions of the subject provoke invariable resistance and conditions in which all desires are immediately satisfied without any effort of the subject In the latter situation, the requirement in search activity does not develop, or may even fade, because such activity simply becomes unnecessary. If the requirement in search activity develops insufficiently, subsequent search can be determined only by means of the situation If a subject with a pronounced search requirement is constantly punished and frustrated by inevitable failures and obstacles, the subject may develop a renunciation of search despite the high level of search requirement So renunciation of search may reflect either a negative feedback between behavior and its outcome or the sequence of the low level of search requirement in situations that do not stimulate subjects to develop search behavior. Two Divergent Forms of Emotional Tension in Humans and Animals Tracing the boundaries of search activity in man is a complex problem, because cognitive processes that are not accompanied by overt behavior play a great role Search activity manifests itself in the planning, imagining, and reappraising of the situation Thus, the concept of search activity covers a broad range of overt as well as covert aspects of behavior Nevertheless, search activity, as defined by Rotenberg (1984), clarifies which behavioral categories cannot belong This applies primarily to all forms of stereotyped behavior with a precise prediction of results, as well as to pathological anxiety and panic behavior At first glance, panic behavior may seem to imitate search behavior, but differs from it by the disturbance of the feedback between the activity and its regulation During panic behavior, the results of activity are not considered at any stage and, therefore, cannot be used for the correction of behavior (e g, changing its direction), hence, no line of activity can be traced to its pragmatic conclusion, making panic behavior easily imitative, approaching stereotyped behavior According to clinical observations, panic behavior during catastrophic events is usually displaced by or combined with depression (Rosenbaum & Seligman, 1989, Zuckerman, 1984) According to Panksepp (1982), the freezing reaction, which he confers to the fear system, also characterizes a panic system In addition, tricyclic antidepressants block panic behavior (Klein, 1981). The cardiac-metabolic dissociation mentioned earlier may be a sign of the pathological-like anxiety that can accompany any form of task solving during renunciation of search The term anxiety, however, is ambiguous There is a distinction between emotional tension that arises in a healthy person during a stressful situation and pathological anxiety (Table 1) The so-called normal anxiety that stimulates exploratory behavior (Halliday, 1966) and reflects the mobilization of all resources to overcome obstacles is opposite to depression in its neurochemical manifestations (Gray, 1982) Pathological anxiety reduces the capability to cope with stress (Lader, 1975, Sloman, 1976), and is usually combined with depression (Roth, 1979) Further more, pathological anxiety is similar to depression in its neurochemical basis (Zuckerman, 1982) Alpert and Haber (1960) developed a psychological test to discriminate between so-called productive and unproductive anxiety, but the validity of this test is questionable (Wittmaier, 1974) The exact physiological correlates of these two types of anxiety are unknown, but there is some evidence for the hypothesis that the high rate of unspecific electrodermal responses (EDRs), the slow habituation of EDRs, and the diminished amplitude of EDR on presentation of the first stimulus may indicate the predominance of an unproductive, pathological anxiety (Lader, 1979) In clinically anxious individuals, indicators of autonomic arousal in response to various stimuli usually increase less than in normal controls (Lader, 1983). Consequently, pathological anxiety and panic behavior can both lead to depression and must be distinguished from the normal emotional tension connected with search activity The difference between pathological anxiety and panic behavior is present in their genesis, because pathological anxiety is due to repression and therefore reflects a state of renunciation of search from the very beginning, whereas panic behavior can be considered an extreme of normal anxiety or emotional tension in a stressful situation, when the motivation to avoid the obstacle is excessively high and search behavior is shifted to renunciation of search. The concept that such global behavioral patterns as search and renunciation of search underlie different definite forms of emotional behavior can help to explain why systems that seem to be opposite (for instance the rage system and the expectation system) may also show synergistic activity under some circumstances (Panksepp, 1982) Freezing is another example of overt behavior that may belong either to search activity or to renunciation of search. Experiments involving the ventrome- dial hypothalamic nucleus in rats and rabbits (Rotenberg & Arshavsky, 1979b), as well as studies of learned helplessness in dogs (Overmier & Seligmann, 1967, Seligman, 1975), clearly have demonstrated freezing to be a surrender reaction, expressing renunciation of search Conversely, freezing can also reflect the temporary blocking of overt behavior while facing an unexpected and significant, or dangerous situation During the absence of motor behavior, the animal searches for a new solution, trying to use effective approaches to avoid possible obstacles Freezing can be a state of intense reappraisal of the situation, in which search activity will not be expressed in overt behavior. The difference between the two kinds of freezing can be observed in reactions to environmental changes. Passive avoidance, which appears during states of search activity, can be changed immediately to active behavior in cases of changing conditions and have no negative consequence for body resistance (Rotenberg, 1984) On the contrary, freezing as a surrender reaction is a form of learned helplessness and does not change until some kind of adaptive mechanism is included (Overmier, 1985, Sehgman, 1975). In his conceptual nervous system, Gray (1982, 1987) described freezing behavior as caused by the action of his Behavior Inhibition System (BIS) Despite overt behavior being inhibited, the animal shows increased attention to the environment Thus, the outcome of the BIS, which is activated under conditions of fear and stress by means of noradrenergic pathways from the locus coeruleus and serotonergic fibers from the raphe nuclei, is behavioral inhibition together with an increment of arousal and increased attention ( Gray, 1982, Figure 1-1) However, Gray's behavioral model does not allow for the distinction between freezing that is dependent on search activity and freezing characterized by the renunciation of search, because the overt behavior may be very similar in both manifestations of internal states. It is necessary, therefore, to extend Gray's (1982) unified BIS concept, taking into account the two divergent forms of emotional tension or anxiety one adaptive, the other maladaptive The Behavioral Activation System (BAS) as formulated by Gray (1970), which he identified in his later work with the limbic reward structures (Olds & Olds, 1965), cannot be regarded as the only conceptual model for search activity, because its outcome is contrary to behavioral inhibition and freezing, which also may be observed during search activity. Additionally, the unified action of benzodiazepines on the BIS, and thus on anxiety, which forms the pharmacological basis of a unified BIS concept, has to be questioned because of their paradoxical actions It is well known that benzodiazepames sometimes enhance and sometimes diminish the motor response to a threatening signal (Gray, 1982) They do not block or even increase active avoid-ance, but they usually block passive avoidance, which is often a sign of renunciation of search (Soubrie, 1982) Sometimes anxiolytics are also beneficial for the treatment of depression (Iversen, 1982). From our point of view, the effect of anxiolytics depends on the nature of anxiety The pathological-like, unproductive anxiety (renunciation of search) is more sensitive to anxiolytics, and, because of the competition between renunciation of search and search behavior, passive avoidance can be replaced by active avoidance We believe that various contradictions, ambiguities, and data instabilities by transition from one experiment to another during behavioral investigation of anxiety may result from the failure to discriminate between these two forms of emotional tension, which differ mainly with respect to the presence or absence of search activity. Autonomic Patterns in the Two Forms of Emotional Tension Autonomic nervous system activity patterns should be typical for both forms of emotional tension Search behavior is usually accompanied by the activation of the sympathetic branch and by peripheral norepinephrine excretion However, the activity of the sympathico-adrenal axis cannot be used to discriminate between search activity and renunciation of search First, renunciation of search in a stress ful situation is accompanied by activating the sympathetic system During the freezing reaction, which may be displayed in this situation, arterial blood pressure is often increased, as it is during the fight/flight reaction However, this increase lasts longer after the stress situation is over, as compared with the fight/flight reaction (Valdman et al, 1979) In addition, an elevated level of catecholamine excretions observable during maladaptive (pathological) emotional tension. Second, some forms of search behavior do not require pronounced activity of the sympathico-adrenal axis and, therefore, do not affect cardiovascular reactions For instance, if attention is outwardly oriented, the heart rate decreases despite the fact that search activity is included The greater the amount of the productive emotional tension combined with arousal in this situation, the more pronounced the phasic heart rate decrease will be (Lacey & Lacey, 1974). There is also a marked difference between OR habituation in healthy versus pathological subjects In healthy subjects, the EDR amplitude during the first two ORs is higher and the habituation is faster (Lader & Wing, 1966, Stern & Janes, 1973) This is easy to explain within our theoretical framework Pathological subjects are overwhelmed by their maladaptive emotional tension, which hyper arouses them This high arousal, however, is not productive, and does not help the subject to overcome real obstacles and to solve tasks (see Figure 1) Moreover, if renunciation of search is in competition with search activity, pathological anxiety blocks the possibility of a task solution that requires search activity. If the spontaneous EDR reflects an inner emotional tension (which is not the result of the concentration and attention to reach the goal but instead makes the subject hypersensitive to all irrelevant stimuli), and the evoked EDR reflects the emotional tension during decision making in the temporal delay (Rotenberg & Vedenyapin, 1985, Spinks, & Siddle, 1983), it can be suggested that a healthy person displays a more intense, although shorter lived, initial reaction to an unexpected stimulus Therefore, the ratio between evoked and spontaneous EDRs (amplitudes and rates) may be useful in differentiating between adaptive and mal-adaptive emotional tension. An alternative explanation of the EDR's lower amplitude to an unexpected stimulus in a pathologically anxious person (as compared with a normal subject) may be the so-called law of initial values (Wilder, 1931) Results of psychophysio-logical experiments correspond to Wilder's law if they are performed with pathological subjects or high sensitizers (eg, Boucsein & Frye, 1974, Weinberger, Schwartz, & Davidson, 1979) Nevertheless, Wilder's law cannot account for various results in this context If the subject is involved in a meaningful activity and is highly motivated, the HR may accelerate, and blood pressure and EDR amplitude may increase, regardless of their initial levels (Keefe & Johnson, 1970, Lovallo, Pincomb, & Wilson, 1986b, O'Gorman & Jamieson, 1977) Based on our theory, results with pathological or overaroused subjects can be reinterpreted as follows If the autonomic activity reflects maladaptive emotional tension, it will be much more difficult for the subject to engage in task performance Renunciation of search, being predominant in those subjects, blocks search activity Therefore, the low autonomic reaction in pathologically anxious persons or sensitizers may not be due to the high initial level, but rather to the inability to accept the challenge of the task that requires search behavior. Attempts to explain autonomic reactions solely through body movements and metabolic processes of the organism are also unsuccessful Heart rate acceleration accompanies some purely intellectual tasks (e g, counting) that don't require body movements (Pribram & McGuinness, 1975). But during certain DRs or during the anticipation of stress, the correspondence between autonomic arousal and metabolic processes disappears, because autonomic arousal exceeds the metabolic demands of the organism (Carrol et al, 1986, Stoney, Langer, & Gelling, 1986) There is some reason to believe that such an excess of metabolic demands precisely characterizes maladaptive emotional tension. Data about autonomic correlations of mental task performance also need re-evaluating The excessive HR acceleration does not reflect a mental effort to solve the task, but rather the emotional tension evoked by the task (It must be kept in mind that every intellectual task is a challenge for self-esteem and can, therefore, provoke a DR and emotional tension) The isometric muscle contractions that accompany mental efforts and cause HR acceleration may be a sign of this covert (inhibited) DR (Diamond & Carver, 1980) Thus, HR acceleration may be a result of body movements during overt defense and orienting behavior, or a result of isometric muscle contraction during covert defense reaction Heart rate acceleration that exceeds the metabolic demands reflects maladaptive emotional tension Attempts to discriminate the physiological concomitants of different emotional expenences also produced many contradictions that may partly be removed using the concept of search activity In an investigation performed by Ax (1953), anger, compared with fear, was accompanied by a more pronounced increase of diastolic blood pressure, an increased number of skin conductance responses, a more pronounced decrease in HR, and increased EMG activity In contrast, fear was accompanied by a more pronounced increase in skin conductance and respiration rate. In a subsequent investigation, Funkenstein, King, and Dorlette (1957) established that the physiological concomitants of anger depend on the behavioral form in which the anger is expressed Subjects directing anger toward the experimenter (anger-out reaction) had less pronounced increases in systolic blood pressure and in heart rate than did subjects in the anxiety condition Subjects expressing self-directed anger (anger-in reaction) differed from subjects in the anxiety condition only m their less pronounced cardiac output increase, whereas anger-out differed from anger-in in the same respect as it did from anxiety. Investigations that disregarded these different forms of anger could not clearly separate fear and anger conditions This was true for cardiovascular responses (Hams, Shoenfeld, Gwynne, Weissler, & Warren, 1964) For example, Stemmler (1984) found that fear was accompanied by increased vasoconstnction, and anger by vasodilation However, Funkenstem (1956), Chessiec, Bassan, and Shattan (1966), Schwartz, Weinberger, and Singer (1981), Roberts and Weerts (1982), as well as James, Yee, Harshfield, Blanke, and Pickering (1986) found anger to be accompanied predominantly by the increase of diastohc blood pressure, caused by increased vasoconstriction Systolic blood pressure, which depends on heart rate and cardiac output, shows higher correlations with anxiety (Funkenstem, 1956) and with suppressed anger (Dimsdale et al, 1986) These results agree with data suggesting that blood pressure caused by cardiac output is controlled by epinephrine, and blood pressure caused by increasing peripheral vasoconstriction is controlled by norepinephrine (Funkenstein, 1956), because active and aggressive emotional display (anger-out) is accompanied by increased norepinephrine excretion with or without epinephrine excretion In contrast, tense and anxious but passive behavior is accompanied by epinephrine excretion (Cohen & Silverman, 1959, Elmadjian, Hope, & Lampson, 1957) Epinephrine injection produced increases in heart rate (Funkenstem et al, 1957), which may be used as an indicator of anxiety and fear reaction Norepinephrine injection produced a pattern similar to the anger-out pattern. These results clearly indicate that anger-in is more different from anger-out than from anxiety and fear The difference between anger-out and anger-in is found in the subjects' ability to fight, at least with the experimenter, and therefore can be compared with search activity. Conversely, subjects experiencing anger-in, which may lead to a decrease of self-esteem and surrender reaction, are more prone to renunciation of search. Hormonal Patterns of the Two Forms of Emotional Tension The difference between the two forms of emotional tension that does or does not include search activity is also displayed in the pattern of excretion of the adreno-medullar and adreno-cortical hormones The dynamic of catecholamine excretion is very significant For instance, a positive adaptive value in rats (survival without signs of distress) is associated both with low-arousal base rates of catecholamines, and, in challenge stress contexts, with fast and strong adrenal-medullary arousal generation (Pfeifer, 1976) Rats with early moderate stressful experience (eg, handling), in comparison with rats without such an experience, have larger adrenal glands and lower catecholamine and cortisol base rates and, in the stressful situation, show reduced cortisol responses and sharp catecholamine spikes (more speed and more pronounced increase from base rates with a rapid return to base rate (Meany et al, 1987) That means that previous moderately stressful experiences trained the physiological coping mechanisms and increased the coping skills in the rats. However, it is also well known that previous moderate stress can immunize subjects against learned helplessness and renunciation of search That is why it is possible to suggest that immunization against renunciation of search itself has a positive effect on the physiological coping mechanisms The results cited here also lead to the conclusion that the ratio of challenge/base rates of catecholamine excretion may reflect the functional sufficiency of coping mechanisms and display the adaptive emotional tension The better the task performance in stressful situations, the larger the adrenaline excretion increase from base rate to task (Ellerstein, Johnsen, & Ursin, 1978, Johansson & Frakenhaeuser, 1973, O'Hanlon & Beatty, 1976, Rauste-von Wright, von Wright, & Frankenhaeuser, 1981) People whose adrenaline levels increased from base rate to task performance had better social adjustment, higher achievement, were more stable and less anxious, and had higher ego strength (Forsman 1981, Rauste-von Wright et al, 1981, Roessler, Burc, & Mefferd, 1967) The interrelations between epinephrine and norepinephrine excretion also seems to be sensitive to the form of behavior in stressful situations and hence to the type of emotional tension If a subject increases his or her ability to avoid an electric shock from one training session to another, but after the final training session is still unable to avoid more than 60% of the shocks (and that is why the forecast of the results of the subject's activity remains indefinite to him or her), the decrease of epinephrine excretion from session to session is accompanied by a stable excretion of norepinephrine (Frankenhauser & Rissler, 1970) Frankenhaeuser explains that the decrease of epinephrine excretion reflects the displacement of helplessness by the ability to master the task, although Wagner (1989) explains the stability of the norepinephrine excretion as a consequence of anger caused by the 40% of failures From our point of view, anger is not necessary in this condition because it may also diminish in tandem with the ability to master the task, at least during the first few sessions But, because of the 40% of failures, the outcome of every subsequent session remains unpredictable for the subject Shock avoidance does not become a stereotyped behavior and such an unpredictable situation stimulates search activity, especially in the context of the previous success. Starting from this point, we can suggest that the above-mentioned correlation between anger-out behavior and norepinephrine excretion (Cohen & Silverman, 1959, Elmadjian et al, 1957) also reflects the component of search activity in anger-out behavior This hypothesis is confirmed by data that demonstrates that norepinephrine excretion followed ratings of concentration and irritation during the process of the solution of complex sensorimotor tasks, whereas epinephrine (E) excretion followed ratings of unpleasantness and irritation (Frankenhaeuser, Nordheden, Myrsten, & Post, 1971) Norepinephrine (NE) excretion was also increased in subjects who had to conduct a difficult sorting task while under the influence of stressors such as loud noise, bright light, and time pressure (Carlson, Levi, & Oro, 1972) The NE/E ratio may be important for the differentiation of the two forms of emotional tension, although further investigations in this direction are essential. The correlation between cortisol and catecholamine excretion also may serve as a useful index Adaptive emotional tension, which corresponds with active coping, is accompanied by the predomination of the catecholamine excretion, whereas maladaptive emotional tension, which reflect distress, is characterized by the increase of cortisol excretion either combined or not combined with catecholamine excretion For instance, paratrooper trainees showed sustained daily-task-related catecholamine increases that were not associated with distress (Baade, Ellerstein, Johnson, & Ursin, 1978) In contrast, high cortisol base rates are associated with depression, anxiety, and neuroticism (Anisman & LaPierre, 1982) Type A's with a family history of cardiovascular disease had higher cortisol levels than did Type As without such history (Ursm & Knardahl, 1985) Both catechola-mines (E and NE) may be excreted during a variety of emotions, particularly when the situation involves uncertainty and lack of control (Frankenhaeuser, 1984). It is important to compare these results with data presented by Williams (1986) Only hostility and anger suppression-but not challenge seeking per se- correlates with coronary heart disease The suppression of hostility and anger represents the renunciation of search because the behavior that accompanies hostility and anger is blocked This suggests that working hard and seeking challenge, which characterize Type A subjects, do not predispose a subject to coronary heart disease, but rather suppressed hostility and anger, often caused by unsatisfied competitive attitudes, are responsible, which characterize only a part of Type A personality It is possible to hypothesize that exactly suppressed hostility and anger correlate with increased cortisol excretion According to our conception, uncertainty can determine search activity, although renunciation of search may also appear in this condition (perhaps, explaining the elevation of both catecholamines) Heart rate and adrenaline were elevated even in experienced air-traffic controllers, but high cortisol responses occurred only in defensive individuals (Ursin, Murison, & Knardahl, 1983) During distress-free efforts, catecholamines increased, but cortisol responses remained below base rate in humans (Frankenhaeuser Lundberg, & Forsman, 1980) In efforts with distress, both adrenaline and cortisol levels were elevated (Lundberg & Frankenhaeuser, 1980). Cortisol elevations in stressful situations are not maintained by the high ac tivity levels per se, but rather by a lack of appropriate responses (Dienstbier, 1989) Cortisol elevation together with catecholamine depletion predominates during unavoidable stress if it causes learned helplessness (renunciation of search) But even the ineffective, albeit active, response of fighting among rats subjected to inescapable shock inhibits catecholamine depletion (Anisman & La-Pierre, 1982) This means that the process, not the activity that includes search behavior, prevents the negative consequences of stress, cortisol elevation may be the sign of the situation subjectively estimated as unavoidable. Underestimation produced by the dull vigilance task causes cortisol elevation, especially in activity-seeking Type A subjects (Lundberg & Forsman, 1979), which means that the decrease of search activity may cause distress, especially in people with a high requirement in search activity Thus, it is possible to use the catecholamine/cortisol ratio for the estimation of the quality of emotional tension. Sleep and the Two Forms of Emotional Tension One of the best indicators of search activity in animals is a high amplitude and well-organized hippocampal theta rhythm All forms of behavior that are accompanied by hippocampal theta (e g, orienting, learning, searching for food or escape, and startle) also include search activity Conversely, theta rhythm is absent during freezing, reflecting the reaction of surrender, or during stereotyped consummatory behavior with a definite predictability of the behavioral outcome (e g, during the performance of an unconditioned reaction) Because hippocampal theta cannot be recorded in humans, the occurrence of search activity during daytime can be recorded by means of sleep structure. Behavioral forms, including a search component, are accompanied by a reduction in REM sleep without a subsequent rebound, meaning there is a drop in the REM sleep requirement Conversely, the state of renunciation of search al most doubles the REM-sleep requirement and the percentage of this sleep phase in rate (Rotenberg & Arshavsky, 1979a, 1979b) REM sleep is a mechanism that permits compensation for the biologically and psychologically harmful state of renunciation of search and ensures the restoration of search activity (Rotenberg, 1984). Proceeding from the search activity concept, (Rotenberg & Arshavsky, 1979a), we analyzed sleep changes in 27 healthy 21- to 30-year-old students who had no complaints about sleep disturbances with respect to their reactions to stress Their night sleep was studied in a laboratory on a post-examination night and on one of the post-holiday nights Thirty min before the examination and 30 mm after, all subjects had their pulse frequency, arterial blood pressure, and bio-electrical activity of the orbicular muscles of the mouth recorded The control data on these indices were obtained on one of the term days (at the same time of day)
Before the examination, all subjects showed a rise in pulse frequency, arterial blood pressure, and bioelectrical activity of the orbicular muscles of the mouth, compared with the control measures, however, after the examination, two groups could be separated The Group 1 subjects (16 students) indices dropped to the control level 30 mm after the examination, whereas the indices of subjects in Group 2 remained at a stable, high level, tending to rise as compared with pre-ex-amination data
The groups exhibited substantial distinctions in their night sleep structure The subjects of Group 2 demonstrated a significant increase in the percentage of REM sleep on the post-examination night compared with the control night and with the subjects of Group 1 on the post-examination night On the post-examination night, the subjects of Group 2 showed a positive correlation between the total duration of sleep and the proportion of REM sleep (+ 74) and a negative correlation (- 6) between the duration of delta sleep and that of REM sleep
We assume that the distinctions between the two groups reflect the distinctions between two types of reactions to emotional stress.
This view is also supported by the fact that the subjects of Group 2 showed worse results in resolving logical problems on the evening after the examination than on the following morning, also, their results were worse than those of Group 1 Additional support comes from data demonstrating that slow post-stress cate-cholamine decline is accompanied by poor performance, neuroticism, and anxiety (Eysenck, 1983, Lader, 1983) Slow post-stress decline of physiological parameters undoubtedly reflects the same processes as slow catecholamine decline. Rotenberg and Arshavsky (1979a) suggest a new indicator for characterizing the quality of pre-sleep emotional tension, which is the ratio of REM sleep to delta sleep in the first two sleep cycles In the case of a pronounced pathological anxiety, a rise in the REM sleep requirement is frequently paralleled by a reduction of the delta sleep in the first two cycles, which has the cumulative effect of increasing this index For instance, in 28 pathological patients with an anxiety syn drome, this index was 0 8(0 08), whereas in 6 healthy individuals with high achievement motivation it was 0 22 (0 1) Night sleep can thus be used as an indicator of the quality of emotional tension and the type of reaction to stress. Rotenberg and Alexeyev (1981) investigated the reactions to stress (emotional and intellectual simultaneously) in 12 subjects suffering from the initial stages of essential hypertension A careful clinical and paraclinical examination delineated the symptomatic character of the disease The examined patients were 25-40 years old (average age 34) Emotional stress was induced along the lines of the classical level-of-aspiration experiment suggested by K. Levin The dis- crimination of the level of aspiration, which was arbitrarily carried out by the experimenter, led to the realization that an objectionably reduced level of the subject's intellectual training created the situation of emotional stress The experiment was preceded and followed by polygraphic registration of HR, skin rheovasography, and measurements of arterial pressure On the nights before and after the emotional stress, polygraphic sleep studies were performed The pre-stress control night was preceded by an adaptation night The patients were examined using the MMPI. In Group 1 patients, emotional stress was followed by longer delta sleep, in Group 2, by REM sleep In both instances, stress led to a rise in arterial pressure, compared with baseline value In Group 1, however, this was due to a rise m the general peripheral vascular resistance with a relative drop in the cardiac output and the blood volume In Group 2, the rise in arterial pressure resulted from an expansion of the cardiac output accompanied by some drop of peripheral vascular resistance It is highly relevant that the rise in arterial pressure was accompanied by a rise in cardiac volume only in Group 2 patients The minute change in volume m Group 1 subjects did not display a significant post-stress change. On the post-stress night, their delta sleep time became longer and their REM sleep decreased Patients of the second group exhibited a longer REM sleep time, a shorter delta sleep time, and a negative REM-delta sleep correlation (r= - 47) A control group of 8 subjects, for whom the third night was registered without previous stress, did not exhibit these sleep structure changes. These two types of sleep change strikingly resemble two types of reactions of healthy students to examination stress These experiments support the hypothesis that an excessive rise in cardiac output in a stress condition that does nor require motor activity may be a feature of unproductive (maladaptive) emotional tension This conclusion concurs with the results of Funkenstein's (1956) investigations He found that patients who showed the profound effect of metacholine on blood pressure expressed depression or anxiety, whereas patients who showed the smaller effect expressed interpersonal anger Metacholine has more sustained effects on the E-produced increased output than on the NE-produced vasoconstriction So blood pressure caused by cardiac output is related to depression or anxiety and may cause a real psychosomatic disturbance Whitehead, Blackwell, De Silva, and Robingson (1977) found that blood pressure in hypertensive patients is more related to anxiety than to anger. Conclusions for Psychophysiological Studies Using human psychophysiological experiments for discriminating the two opposite states of emotional tension based on the concept that search activity, as opposed to renunciation of search, requires specific methodological approaches, independent criteria for indicating the presence of either search activity or renunciation of search are required These may be taken from the three levels of psychophysiological recording during an experimental stress situation (a) the subjectively recorded attitude toward the outcome (e g, the tendency to give up and to terminate activity in case of renunciation of search), (b) the measurable behavioral outcome of this activity (e g, the task performance, which should be considerably higher during search activity), and (c) as a first approach to physiological level, the sleep structure may be used as a criterion, especially the proposed REM-to-delta ratio taken from the first two sleep cycles If this ratio is below 0 3, this indicates a prevalence of adaptive emotional tension, whereas ratios exceeding 0 7 point indicate a predominance of maladaptive emotional tension instead In addition, it will be necessary to record the following complex set of physiological parameters and to analyze their interrelationships as well as their dynamics as transients while turning from one task to another 1. The estimation of the interrelations between cardiac output and peripheral resistance of blood vessels in the process of mental task solution It can be assumed that adaptive emotional tension is characterized by the prevalence of the peripheral resistance of blood vessels, whereas cardiac output is prevalent during maladaptive emotional tension
It should be kept in mind that the tasks used to induce opposite emotional states must differ in both quality and difficulty to ensure coverage of the required range of emotional tension All of these propositions are based on the fundamental assumption that the main difference between adaptive and maladaptive emotional tension is determined by the representation of search activity in behavior accompanied by emotional tension This point of view may be helpful not only for the distinction between the two forms of emotional tension but also for the solution of some contradictions present in the psychophysiological literature. REFERENCES Alpert, R, & Haber, R (1960) Anxiety in academic achievement situations Journal of Abnormal and Social Ps\chology, 61, 207-215 Anderson, D E, & Jingling, J E (1978) Total peripheral resistance changes in dogs during aversive classical conditioning The Pavlovian Journal of Biological Sciences, 13, 241-246 Anisman, H, & LaPierre, Y (1982) Neurochemical aspects of stress and depression Formulations and caveats In R W Neufeld (Ed ), Psychological stress and psychopathol-ogy(pp 179-217) New York McGraw-Hill Atkmson, J W (1974) Strength of motivation and efficiency of performance In J W Atkmson & J Raynor (Eds ), Motivation and achievement Washington, DC Winston Ax, A F (1953) The physiological differentiation between fear and anger in humans Psychosomatic Medicine, 15, 433-442 Baade, E, Ellerstein, B, Johnsen, T B, & Ursin, H (1978) Physiology, psychology and performance In H Ursm, E Baade, &S Levine (Eds ), Psychobiology of stress A study of coping men (pp 163-182) San Diego Academic Press. Boucsein, W, & Frye, M (1974) Physiologische und psychische wirkungen von miberfol-gsstress unter berucksichtigung des merkmals repression-sensitization [The physiological and psychic effect ot stress of failure in the context of repression-sensitization] Zeitschrift fur Experimentelle und Angewandte Psychologic, 21 339-366. Carlson, L A, Levi, L, & Oro, L (1972) Stressor-induced changes in plasma lipids and urinary excretion of catecholamines and their modification by nicotinic acid In L Levi (Ed ), Stress and distress in response to psychosocial stimuli Acta medica Scandinavica (Vol Supplementum 528) Stockholm Almquist & Wicksell Carrol, D, Turner, I R, & Hellawell, J C (1986) Heart rate and oxygen consumption during active psychophysiological challenge The effects of level of difficulty Psycho-physiology, 23, 174-181 Chessiec, R D, Bassan, M, & Shattan, S (1966) A comparison of the effect of infused catecholammes and certain affect states American Journal of Psychiatry 123, 156-165 Chodoff, P (1970) The German concentration camps as A psychological stress Archiv General Psychiatry, 22 78-87 Cohen, S I, & Silverman, A J (1959) Psychophysiological investigations of vascular response variability Journal of Psychosomatic Research, 3, 185-210 Diamond, E, & Carver, C (1980) Sensory processing, cardiovascular reactivity and the type A coronary-prone behavior pattern Biological Psychology, 10, 265-275 Dienstbier, R A (1989) Arousal and physiological toughness Implications for mental and physical health Psychological Review, 96, 93 Dimsdale, J E, Pierce, C, Schoenfeld, D, Brown, A, Zusman, R, & Graham, R (1986) Suppressed anger and blood pressure The effects of race, sex, social class, obesity, and age Psychosomatic Medicine, 48, 430-436 Ellerstein, B, Johnsen, T B, & Ursin, H (1978) Relationship between the hormonal responses to activation and coping In H Ursin, E Baade, & S Levine (Eds ), Psychobi ology of stress A study of coping men (pp 105-124) San Diego Academic Press Elmadjian, F J, Hope, M, & Lampson, E T (1957) Excretion of E and NE in various emotional states Journal of Clinical Endocrinology, 17, 608-620 Eysenck, H J (1983) Psychophysiology and personality Extraversion, neuroticism, and psychoticism In A Gale & J A Edwards (Eds), Physiological correlates of human behavior Vol HI Individual differences and psychopatholog\ (pp 13-30) London Academic Press Forsman, L (1981) Habitual catecholamine excretion and its relation to habitual distress Biological Psychology, 11, 83-97 Frankenhaeuser, M (1984) Psychoneuroendocrine approaches to the study of stressful person-environment transactions In H Selye (Ed ), Selye's Guide to Stress Research (pp 46-70) New York Van Nostrand Reinhold Frankenhaeuser, M, Lundberg, U, & Forsman, L (1980) Dissociation between sympathetic-adrenal and pituitary-adrenal responses to an achievement situation characterized by high controllability Comparison between Type A and Type B males and females Biological Psychology, 10, 79-91 Frankenhaeuser, M, Nordheden, B, Myrsten, A L, & Post, B (1971) Psychophysiological reactions to understimulation and overstimulation Acta Psychologica, 35, 298-308 Frankenhaeuser, M, & Rissler, A (1970) Effects of punishment on catecholamme release and efficiency of performance Psychopharmacologia, 17, 378-390 Funkenstein, D H (1956) Nor-E-like and E-like substances in relation to human behavior Journal of Nervous and Mental Diseases, 124, 58-68 Funkenstem, D H, King, S H, & Dorlette, M (1957) Mastery of stress Cambridge, MA Harvard University Press Gal, R, & Lazarus, R S (1975) The role of activity in anticipating and confronting stressful situations Journal of Human Stress, 1, 4-20 dinner, J A, Bedi, J F, & Horvath, S M (1979) Somatic and non-somatic influences on the heart Hemodynamic changes Psychophysiology, 16 358-362 Goesling, W J, Buchholz, A R, & Carreira, C J (1974) Conditioned immobility and ulcer development in rats Journal of General Psychology, 91, 231-236 Graham, F K (1979) Distinguishing among orienting, defense, and startle reflexes In H D Kimmel, E H van Olst, & J F Orlebeke (Eds ), The orienting reflex in humans (pp 137-167) Hillsdale, NJ Erlbaum Grastyan, E (1976) Motivation and reinforcement Acta Physiologica Academiae Scienti-arum Hungaricae, 48(4), 299-322 Gray, J A (1970) The psychophysiological basis of introversion-extraversion Behaviour, Research, and Therapy, 8, 249-266 Gray, J A (1982) The neuropsychology of anxiety An inquiry into the functions of the septo-hippocampal system Oxford, England Clarendon Press Gray, J A (1987) The psychology of fear and stress Cambridge, England Cambridge University Press Harburg, E, Blakelock, E H, & Roeper, P (1979) Resentful and reflective coping with arbitrary authority and blood pressure Psychosomatic Medicine, 41, 189-202 Halliday, M S (1966) Exploration and fear in the rat Symposia of the Zoological Society of London, 18, 45-59 Harris, W S, Shoenfeld, C D, Gwynne, P H, Weissler, A M, & Warren, J V (1964. Circulatory and humoral responses to fear and anger Journal of Laboratory and Clinical Medicine, 64, 867 (abstract) Iversen, S D (1982) Integrating the literature of anxiety, memory and the hippocampus Commentary to Gray, J.: Neuropsychology of anxiety Behavioral and Brain Sciences,3, 487-488 James, G D, Yee, L S, Harshfield, G A, Blanke, S G, & Pickering, T (1986) The influence of happiness, anger, and anxiety on the blood pressure of borderline hypertensives Psychosomatic Medicine, 48, 502-508 James, S A, Hartnett, S A, & Kalsbeek, W D (1983) John Henryism and blood pressure difference among men Journal of Behavioral Medicine, 6, 259-278. Johansson, G, & Frankenhaeuser, J (1973) Temporal factors in symphato-adrenomedullary activity following acute behavioral activation Biological Psychology,I, 63-73 Keefe, F B, & Johnson, L C (1970) Cardiovascular responses to auditory stimuli Psychonomic Science, 19, 335-337. Klein, D F (1981) Anxiety reconceptualized In D F Klein & J Rabkin (Eds ), Anxiety New research and changing concepts (pp 235-263) New York Raven Press. Lacey, B C, & Lacey, J I (1974) Studies of heart rate and other bodily processes insensorimotor behavior In P A Obrist, A H Black, J Brener, & L V DiCara (Eds ), Cardiovascular psychophysiology Current issues in response mechanisms Chicago: Aldme-Atherton. Lacey, J I (1967) Somatic response patterning and stress Some revisions of activation theory In M H Appley & R Trumbull (Eds ), Psychological stress Issues in research (pp 14-37) New York: Appleton. Lacey, J I, & Lacey, B C (1970) Some autonomic-central nervous system interrelationships. In P Black (Ed ), Physiological correlates of emotion San Diego: Academic Press Lader, M (1975) The psychophysiology of mental illness. London and Boston: Routledge & Kegan Paul Lader, M (1979) The orienting reflex in anxiety and schizophrenia In H D Kimmel, E H van Olst, & J F Orlebeke (Eds. ), The orienting reflex in humans (pp 607-617) Hillsdale, NJ Erlbaum . Lader, M (1983) Anxiety and depression In A Gale& J A Edwards (Eds ), Physiological correlates of human behavior Vol III Individual differences and psychopathology (pp 155-167) London Academic Press Lader, N, & Wing, L (1966) Physiological measures, sedative drugs and morbid anxiety (Vol 14) In Institute of Psychiatry, Maudsley Monographs (Ed ), London Oxford University Press Lens, W, & DeVoider, M (1980) Achievement motivation and intelligence test scores. A test of the Yerkes-Dodson hypothesis Psychol Belg 49-59 Lovallo, W R, Pincomb, G A, & Wilson, M F (1986a) Heart rate activity and Type A behavior as modifiers of physiological response to active and passive coping Psycho-physiology 23, 105-112. Lovallo, W R, Pmcomb, G A, & Wilson, M F (1986b) Predicting response to A reaction time task Heart rate reactivity compared with Type A behavior Psychophysiology, 23 648-656. Lundberg, U, & Foreman, L (1979) Adrenal-medullary and adrenal-cortical responses to understimulation and overstimulation Comparison between Type A and Type B persons Biological Psychology, 9, 79-89. Lundberg, U, & Frankenhaeuser, M (1980) Pituitary-adrenal and sympathetic-adrenal correlates of distress and effort Journal of Psychosomatic Research, 24 125-130. Meany, M J, Aitkens, D H, Berkel, C, Bhatnagar, S, Sarrieau, A, & Sopolsky, R M (1987) Post-natal handling attenuates age related changes the adrenocortical stress response and spatial memory deficits the rat Paper presented at the 17th Annual Meeting of the Society of Neuroscience, New Orleans, LA Obrist, P A, Gaebelin, C J, Teller, E S, Langer, A W, Gringnolo, A, Kathleen, C L, & McCubbin, J A (1978) The relationship among heart rate, carotid dP/dt and blood pressure and myocardial performance Psychophysiology, 15, 292-301. Obnst, P A, Light, K C, McCubbin, J A, Hutcheson, J S, & Hoffer, J L (1979) Pulse transit time Relationship to blood pressure and myocardial performance Psychophysiology, 16, 292-301. O'Gorman, J G, & Jamieson, R D (1977) Short latency acceleration of human heart rate as A function of stimulus intensity Perceptual and Motor Skills, 45, 579-583. O'Hanlon, J F, & Beatty, J (1976) Catecholamine correlates of radar monitoring performance Biological Psychology, 4, 293-304. Olds, J, & Olds, M E (1965) Drives, rewards, and the brain In T M Newcomb (Ed ), New directions in psychology II New York Holt Overmier, J B (1985) Toward A reanalysis of the causal structure of the learned helplessness syndrome In F R Brush & J B Overmier (Eds ), Affect, conditioning and cognition Essays on the determinants of behavior (pp 211-227) Hillsdale, NJ Erl-baum. Overmier, J B, & Seligman, (1967) Effects of unescapable shocks upon subsequent escape and avoidance responding Journal of Comparative and Physiological Psychology, 63, 28-33. Panksepp, J (1982) Toward A general psychobiological theory of emotions The Beha\ loral and Brain Sciences, 5 407-467 Pfeifer, W D (1976) Modification of adrenal tyrosine hydroxylase activity in rats following manipulation in infancy In E Usdin, R Kvetnansky & I J Kopin (Eds ), Catechola mines and stress (pp 265-270) Oxford, England Pergamon Pribram, K H, & McGuinness, D (1975) Arousal, activation and effort in the control of attention Psychological Review, 82, 116-149 Ramirez, A, & Lasater, T (1976) Attitudmal and behavioral reactions to fear-arousing communications (Vols 811-817) In Psychological Review (Ed ), 38. Rauste-von Wright, M, von Wright, J, & Frankenhaeuser, M (1981) Relationships between sex-related psychological characteristics during adolescence and catecholamine excretion during achievement stress Psychophysiology, 18, 362-370. Roberts, R J, & Weerts, T C (1982) Cardiovascular responding during anger and fear imagery Psychological Reports 50, 219-230. Roessler, R, Burc, N R, & Mefferd, R B (1967) Personality correlates of catecholamme excretion under stress Journal of Psychosomatic Research, 11, 181-185. Rosenbaum, D L, & Seligman, M E P (1989) Abnormal psychology New York Norton Rotenberg, V S (1984) Search activity in the context of psychosomatic disturbances, sleep of brain monoamines and REM function Pavlovian Journal of Biological Science 19, 1-15. Rotenberg, V S, & Alexeyev, V V (1981) Essential hypertension A psycho-somatic feature or A psycho-somatic disease? A differential analysis of cases in terms of search activity concept Dynamic Psychiatry, 68, 129-140. Rotenberg, V S, & Arshavsky, V V (1979a) REM sleep, stress, and search activity Waking and Sleeping, 3, 235-244 Rotenberg, V S, & Arshavsky, V V (1979b) Search activity and its impact on experimental and clinical pathology Activitas Nervosa Superior (Praha), 21, 105-115. Rotenberg, V S, & Vedenyapin, B (1985) GSR as reflection of decision-making under conditions of delay Pavlolian Journal of Biological Science, 20, 11-14 Roth, M A (1979) A classification of affective disorders based on A synthesis of new and old concepts InE Meyer&J V Brady (Eds ), Research in the psychobiology of human behavior (pp 75-114) Baltimore Johns Hopkins University Press. Schiff, B B, Rusak, B, & Block, R (1971) The determination of reinforcing intracramal stimulation An ecological approach Psychology and Behavior, 7(2), 215-220 Schwartz, G E, Weinberger, D A, & Singer, J A (1981) Cardiovascular differentiation of happiness, sadness, anger and fear following imagery and exercise Psychosomatic Medicine, 43, 343-364 Seligman, M E P (1975) Helplessness On depression, development and death San Francisco Freeman Seliman, M E P (1982) Learned helplessness and life span development Papers presented at the International Conference of Life-Course Research on Human Development Max Planck Institute, West Berlin Sherwood, A, Alien, M T, & Obrist, P A (1986) Evaluation of beta adrenergic influences on cardiovascular and metabolic adjustments of physical and psychological stress Psychophysiology, 23, 89-104 Sloman, L (1976) The role of neurosis in phylogenetic adaptation with particular reference to early man American Journal of Psychiatry 133, 543-547. Sokolov, E N (1963) Perception and the conditioned reflex Oxford, England Perga-mon Press. Soubrie, P (1982) Infemng anxiety and antianxiety effects in animals Behavioral and Brain Science, 502-503. Spielberger, C D (1962) The effects of manifest anxiety on the academic achievement of college students Mental Hygiene, 46, 420-426 Spinks, J A, &Siddle, D (1983) The functional significance of the orienting response In D Siddle (Ed ), Orienting and habituation Perspectives in human research Chichester, England Wiley Stemmler, G (1984) Psychophysiologische Emotwnsmuster Frankfurt, Germany Lang. Stern, J A, & Janes, C L (1973) Personality and psychopathology In W F Prokasy & D C Raskin (Eds ), Electrodermal activity in psychologtcal research San Diego Academic Press. Stoney, C M, Langer, A W, & Gelling, P D (1986) The effect of menstrual cycle phase on cardiovascular and pulmonary responses to behavioral and exercise stress Psycho physiology, 23, 393-402. Stoney, C M, Langer, A W, Sutterer, J R, & Gelling, P D (1987) A comparison of bioteedback assisted cardiodeceleration in Type A and B men Modification of stress-associated cardiopulmonary and hemodynamic adjustments Psychosomatic Medicine, 49 79-87. Ursin, H, & Knardahl, S (1985) Personality factors, neuroendocrine response patterns, and cardiovascular pathology In J F Orlebeke, G Mulder, & L J P van Door-nen (Eds ), Psychophysiology of cardiovascular control (pp 715-731) New York Plenum Ursin, H, Murison, R, & Knardahl, S (1983) Conclusion Sustained activation and disease In H Ursin & R Murison (Eds ), Biological and psychological basis of psychosomatic disease (pp 269-277) Oxford, England Pergamon. Vainstein, J J, & Simonov, P V (1979) Emotional structures of brain and heart Moscow Nauka Publishers. Valdman, A V, Kozlovskaya, M M, & Medvedev, S (1979) Regulation of emotional stress by pharmacological agents (in Russian) Moscow Meditsina Publishers. Wagner, H (1989) The peripheral physiological differentiation of emotions In H. Wagner & A Manstead (Eds ), Handbook of social psichophysiology (pp 78-98) New York Wiley. Wauquier, A (1980) The pharmacology of catecholamme involvement m the neural mechanisms of reward Ada Neurobiologiae Experimentalis, 40, 665-686. Weinberger, D A, Schwartz, G E, & Davidson, R J (1979) Low-anxious, high-anxious, and repressive coping styles Psychometric patterns and behavioral and physiological responses to stress Journal of Abnormal Psychology, 88, 369-380 Weiss, J A (1977) Psychological and behavioral influences of gastrointestinal lesions in animal models In J Maseer & M Seligman (Eds), Psychopathology Experimental models (pp 232-269) San Francisco Freeman Whitehead, W E, Blackwell, B, DeSilva, H, & Robingson, A (1977) Anxiety and anger in hypertension Journal of Psychosomatic Research, 21, 383-389 Wilder, J (1931) Das 'Ausgangswert-Gesetz"-em unbeachtetes biologisches Gesetz, seine Bedeutung fur Forschung und Praxis Klinische Wochenschrift, 41, 1889-1893. Williams, R B, Jr (1986) Beyond Type A Psychological and physiological refinements. Paper presented at the annual meeting of the American Psychological Association, Washington, DC Wittmaier, B C (1974) Test anxiety, mood and performance Journal of Personality and Social Psychology 29, 664-669. Yerkes, R M, & Dodson, J D (1908) The relation of strength of stimulus to rapidity of habit formation Journal of Comparative and Neurological Psychology, 18, 459-482 . Zuckerman, M (1982) Leaping up the phylogenetic scale in explaining anxiety Commentary to Gray, J Neuropsychology of anxiety The Behavioral and Brain Sciences, 3, 505-506. Zuckerman, M (1984) Sensation seeking A comparative approach to human traits The Behavioral and Brain Science, 7, 413-434 |